


蜘蛛丝是一种天然的高分子动物蛋白纤维,是蜘蛛4亿年进化的结果[1, 2]。蜘蛛丝的适应进化和生物学功能使其具有独特的物理化学结构和性能[3~5]。蜘蛛丝(特别是大壶状腺丝)具有优良的机械性能(如高强度、弹性、柔韧性和抗断裂性能等)和比重小、耐紫外线、吸能、生物相容性好、生物可降解等特性,其优良的综合性能和特性优于蚕丝等天然蛋白纤维与合成纤维[4~7],可广泛应用于医药、组织工程材料、新型特种纤维、国防、航天航空和日常生活等领域[8]。现已发现并命名的蜘蛛有50, 000种(World spider catalog version 23.0),分布极广,可在几个小时内耐受超过30℃和70%相对湿度的变化幅度。为适应各种变化幅度极大的生态环境,蜘蛛丝纤维的组成、结构、性能与生物学功能等均呈现出种间和种内的差异与多样性[9~13]。因此,蜘蛛丝是一种研究纤维材料结构、性能与功能之间关系的良好模式纤维[14]。人们认识和掌握蜘蛛丝的结构、性能与生物学功能之间的关系,有助于模仿蜘蛛的液晶纺丝工艺与蛛丝特有的物理化学结构和性能以制造符合人们需求的功能纤维[10, 15~17]
影响蜘蛛大壶状腺丝力学性能的因素有很多,如温湿度、纺丝速度、环境介质、氨基酸与丝心蛋白的种类和组成、蜘蛛的大小体重和年龄、蜘蛛捕食猎物的种类大小和体重、卵袋的大小和重量、蜘蛛的营养水平和饥饿程度、纺出来被放置的时间等[7~13]。在自然界中蜘蛛丝会被经常反复拉伸,其反复拉伸特性与力学性能之间有密切的关系[18~21]。但蜘蛛丝反复拉伸特性的材料进化与生态学意义还不十分清楚,测试和验证在生态和演化背景下有关蜘蛛丝特性及其变异的假说是这个领域的研究热点和难点[14]。本文在前期研究结果的基础上[19, 22~26],使用METS (E44.104, USA)和Instron5966(USA)电子万能试验机与Renishaw (Invia Qontor, GB)激光拉曼光谱仪测试蜘蛛大壶状腺丝的定间隔时间且定伸长长度逐渐增加的反复拉伸力学行为,研究反复拉伸后蛋白二级结构的变化,以模拟大壶状腺丝在自然界中被蜘蛛使用时反复拉伸的情形并分析探讨蜘蛛大壶状腺丝在被反复拉伸的过程中其形变、力学行为与结构的变化及其相互之间的关系,以验证我们对蜘蛛丝反复拉伸特性是蜘蛛对其丝纤维力学性能的一种后加工调节方式的推测。为如人工拉伸蜘蛛丝腺体制备新型功能性纤维[31, 32]的仿生设计提供科学依据与理论支撑。
1 实验方法
实验用材料为横纹金蛛(Argiope bruennichi)纺自大壶状腺的卵袋框丝。使用METS电子万能试验机测试蜘蛛丝的定伸长反复拉伸力学行为,定伸长长度增幅为0.5 mm或者1.0 mm,每组内丝样品定伸长反复拉伸5次,两次拉伸的间隔时间为15~25 s或5 min,每组之间的间隔时间≥1 h,共重复7个丝样品。对在空气中7个丝样品的反复拉伸实验做了以下交替设计:(1)组间长时间间隔(≥2 h),组内拉伸间隔为5 min,伸长长度不变或增加;(2)组内组间间隔时间均为5 min,伸长长度增加;(3)组间组内连续拉伸(拉伸间隔大约15~25 s,为测试软件进入下次拉伸点击操作所需的时间),伸长长度增加;(4)长时间间隔(≥1 h),组内连续拉伸(拉伸间隔大约15~25 s),伸长长度增加或不变。测试条件:制作20 mm纸框架,将丝黏贴于其上;夹持距为20 mm,下降速度为10 mm/min,温度为20℃,湿度为65%[19~29]。用SEM观察测量与丝样品相邻近的丝以得到丝纤维的最初直径。由于蛛丝在拉伸过程中体积不变,研究丝纤维的力学性能时使用π(D0/2)2L0=π(D/2)2L[19~29]公式把相对应的负载-位移曲线转换成应力-应变曲线,以得到蛛丝的力学性能参数。使用Spass15.0进行差异性检验分析参数数据。
2 实验结果
2.1 蜘蛛大壶状腺丝反复拉伸力学行为
典型大壶状腺丝由两股细丝纤维构成(图1)。实验结果表明,蜘蛛大壶状腺丝能反复拉伸几十次至一百多次(图2),具有良好的反复拉伸特性。无论是一次拉伸断裂的对照丝样品还是定伸长反复拉伸的丝样品都具有典型大壶状腺丝的力学行为,其负载-位移曲线都比较复杂:由初始弹性区、屈服点、屈服区和加强区及断裂点或终止点组成。其屈服伸长为0.4~0.7 mm,拉伸应变为2.2%~2.6%时发生第二类屈服,曲线出现明显的转折,拉过屈服点后模量骤降,然后应力与模量都趋于平缓,最后进入加强区时应力与模量逐渐增加,出现大壶状腺丝典型的成颈现象。随着拉伸次数与定伸长长度的增加,其力学行为的屈服点负荷也逐渐下降,但是其屈服强度基本保持不变,而初始模量逐渐增加(图2和图3),即随着蛛丝不断反复拉伸其屈服强度基本恒定不变的同时蛛丝变硬(图2)。随着拉伸次数和定伸长长度的增加,蛛丝每次拉伸前的原长呈总体增加的趋势,每次拉伸前的直径和横切面积总体逐渐减小,但是定伸长或定伸长增加幅度很小时其拉伸前的原长变化差异不显著(P>0.05);而经长时间间隔后蛛丝拉伸前的原长比之前稍有缩短(图4)(P<0.05),即蛛丝抗拒被拉长。但是,随着拉伸次数和定伸长长度的继续增加,蛛丝拉伸前的原长变长(P<0.05),即最终拉长直至最后一次拉伸断裂。大壶状腺丝在定伸长长度逐渐增加反复拉伸至一定次数后会先断裂其两股中的一股,断裂成一股后其屈服强度与初始模量是两股的大约2倍(图3)。随着拉伸次数的增加,蛛丝吸收的能量总体趋势是逐渐被耗散一部分,但经过长时间间隔后,其吸收的能量不但没有被耗散反而还增加了,也有不少前后两次拉伸的SS曲线基本重叠的,这时其耗散的能量值几乎接近于零(图3)。
图1

图1
横纹金蛛纺自大壶状腺的金黄色卵袋框丝扫描电镜照片
Fig.1
Scanning electron micrograph illustrating the morphology of typical Mas fiber (diameter is about 20 μm) consisting of double filaments from golden scaffolding silk of A. bruennichi egg case
图2
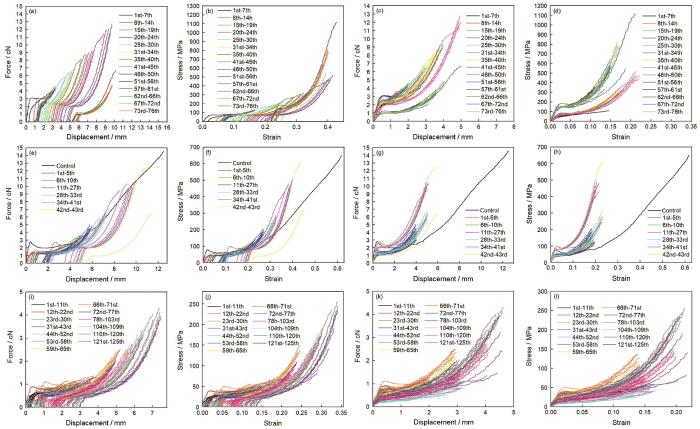
图2
典型性蜘蛛大壶状腺丝反复拉伸力学行为
Fig.2
Typical repeated stretching tensile properties of spider Mas in air. The three silk samples were repeatedly stretched. The curves (a~d, e~h and i~l) are respectively from same silk samples, the curves (b, f and j) are stress-strain curves corresponding to the primitive force-displacement curves (a, e and i) on the testing software. The curves (d, h and l) are true stress-true strain curves corresponding to the force-displacement curves (c, g and k). One of double filaments was broken after a certain number of repeated stretching times
图3
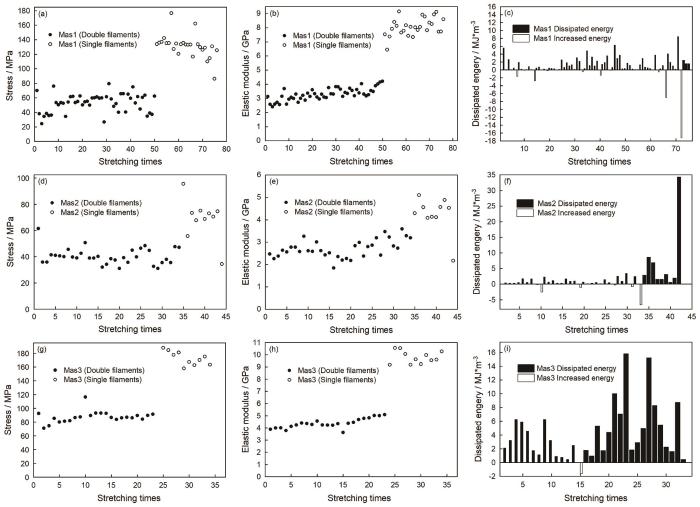
图3
蜘蛛大壶状腺丝反复拉伸过程中典型的力学性能参数的变化
Fig.3
The typical changes of typical mechanical properties parameters during repeated stretching of spider Mas. A~c, d~f and g~i come from the same sample, (a, d and g) yield stress (b, e and h) elastic modulus (c, f and i) dissipated and increased energy, respectively
图4
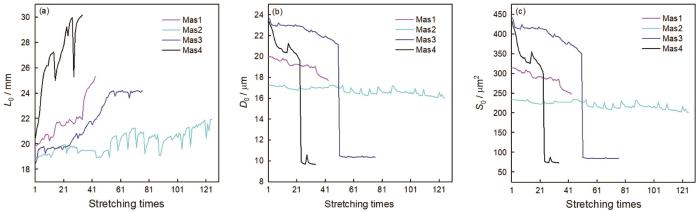
图4
蜘蛛大壶状腺丝反复拉伸次数与其拉伸前的几何参数
Fig.4
Relationship of the geometric parameter before stretching (a) primitive length, (b) primitive diameter, (c) primitive cross sectional area and stretching times of spider Mas. There is a special situation that the initial length was decreased after long interval between two stretching times
2.2 大壶状腺丝的力学行为记忆能力
在定伸长长度增加幅度不大的情况下,组间组内无论间隔5 min(图5)还是连续拉伸即拉伸间隔约为15~25 s(图6),前后两组的力学行为曲线都较好地重叠,即使定伸长长度增加第一次拉伸力学行为曲线与前一组的力学行为曲线的加强区重叠较差而有较小的偏移,但是随后的力学行为曲线的重复性仍然很好(图5c、d、g、h),有时甚至与前一组的力学行为曲线很好地重叠(图5k、l和图6a、b);在定伸长长度不变或增加的情况下,组内两次拉伸之间无论是间隔5 min(图7)还是连续拉伸即拉伸间隔为15~25 s(图6),且经过组间长时间间隔(≥1 h)后只需一次拉伸前后两组力学行为曲线就重叠很好,即只需经过一次拉伸就能不受拉伸历史的影响重现之前的力学行为。前后两次拉伸间隔不同时间梯度的力学行为曲线,表现为一部分不受拉伸历史的影响重现之前的力学行为,另一部分则不能重现之前的力学行为(图8)。天然蜘蛛大壶状腺丝整体表现出良好的力学行为记忆能力。用于激光拉曼光谱测试分析的丝样品总共反复拉伸了34次,其力学行为特征和记忆能力与其它丝样品相同(图9)。
图5
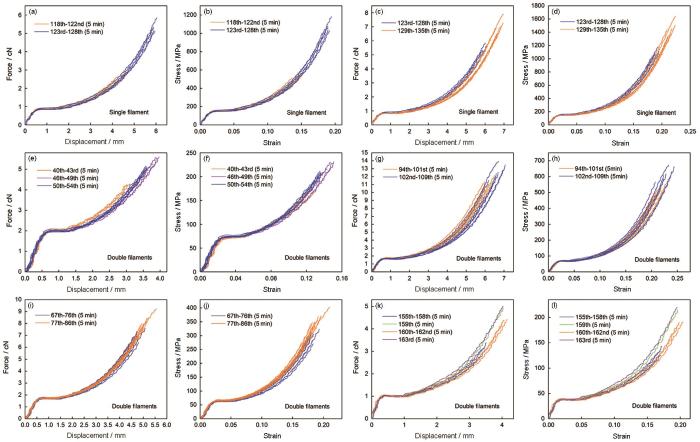
图5
蜘蛛大壶状腺丝定伸长长度增加前后力学行为的比较
Fig.5
Comparison of tensile properties of spider Mas in air before and after increasing fixed elongation. The two silk samples were repeatedly stretched. The curves (a~j and k~l) are respectively from same silk samples. The curves (b, d, f, h, j and l) are true stress-true strain curves corresponding to the force-displacement curves (a, c, e, g, i and k). There is 5 min interval between each stretching. There was significant difference in the original length (L0) of silk samples before stretching between the two groups for Fig.c, d, g, h and i, j (P < 0.05), and the other differences were all not significant (P > 0.05)
图6

图6
蜘蛛大壶状腺丝连续拉伸即很短时间间隔前后力学行为的比较
Fig.6
Continuous stretching tensile properties of the Mas in air. The two silk samples were repeatedly stretched. The curves (a~f and g~l) are respectively from same silk samples. The curves (b, d, f, h, j and l) are true stress-true strain curves corresponding to the force-displacement curves (a, c, e, g, i and k). There is very shot interval about 15 s or 25 s between each stretching except for few stretching. There was significant difference in the original length (L0) of silk samples before stretching between the two groups for Fig.k and l (P < 0.05), and the other differences were all not significant (P > 0.05)
图7
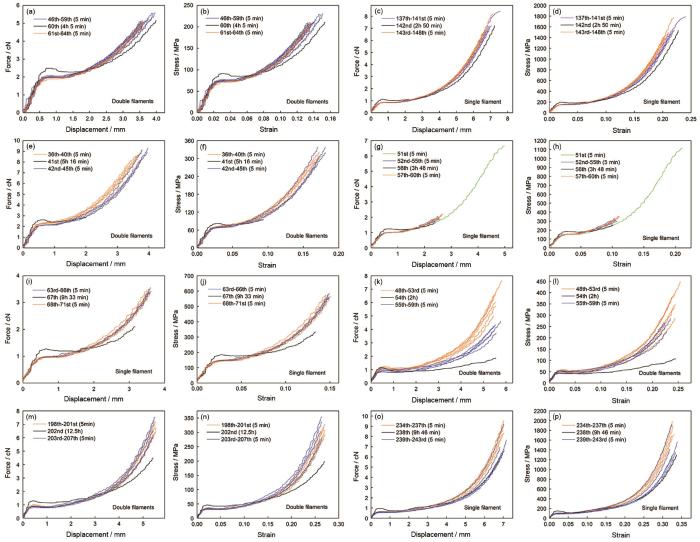
图7
蜘蛛大壶状腺丝力学行为记忆能力
Fig.7
Memory capacity on tensile behavior of spider Mas. The curves (a~d, e~j,k~l and m~p) are respectively from same silk samples. The curves (b, d, f, h, j, l and p) are true stress-true strain curves corresponding to the force-displacement curves (a, c, e, g, i, k and o). These curves show that Mas revert independently from its previous loading history by an about 5 min interval after last subjected to yield and hardening region, furthermore, Mas can reproduce the last time tensile behavior via only one stretch after a few hours or overnight interval. There was significant difference in the original length (L0) of silk samples before stretching between the two groups for Fig.a, b, c, d, k and l (P < 0.05), and the other differences were all not significant (P > 0.05)
图8
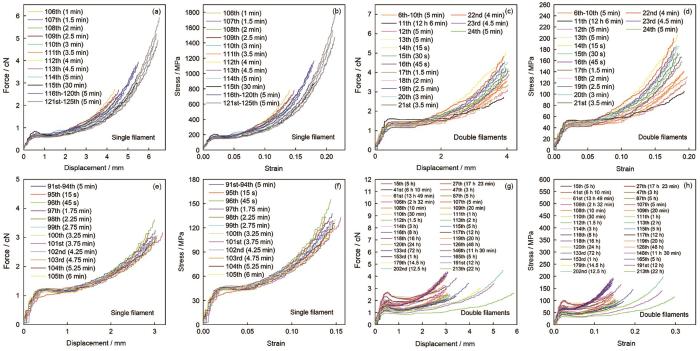
图8
蜘蛛大壶状腺丝间隔不同时间梯度力学行为的比较
Fig.8
Tensile behavior of spider Mas after gradient interval time. The three silk samples were repeatedly stretched. The curves (a~b, c~d, e~h) are respectively from same silk samples. The curves (b, d, f and h) are true stress-true strain curves corresponding to the force-displacement curves (a, c, e and g)
图9
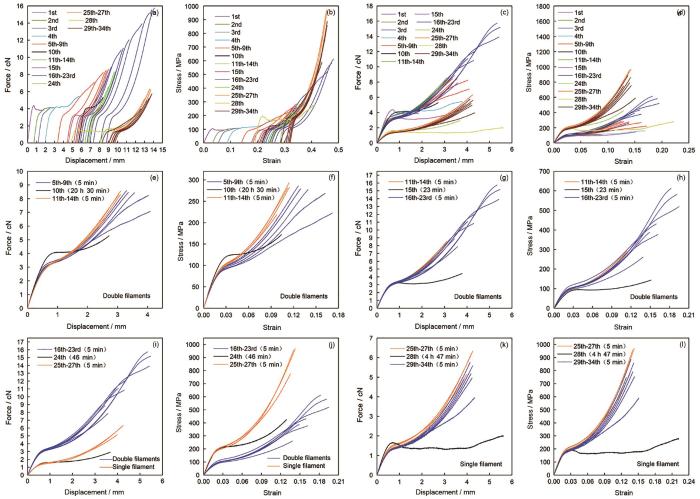
图9
蜘蛛大壶状腺丝定伸长反复拉伸力学行为(用于激光拉曼光谱测试分析)
Fig.9
Repeated tensile behavior of the Mas in air for Raman test. The curves (b, d, f, h, j and l) are true stress-true strain curves corresponding to the force-displacement curves (a, c, e, g, i and k). There was significant difference in the original length (L0) of silk samples before stretching between the two groups for Fig.e and f (P< 0.05), and the other differences were all not significant (P > 0.05)
2.3 激光拉曼光谱分析
图10

图10
蜘蛛大壶状腺丝经不同次数的反复拉伸以及每次拉伸之间不同间隔时间后的激光拉曼光谱(Mas-9th-5 min表示大壶状腺丝被反复拉伸第9次卸载后5 min做的拉曼光谱,其余以此类推)
Fig.10
Raman spectrum of major ampullate gland silk after different stretching times and interval time. (a) mas with double filaments; (b) mas with single filament (except Mas-0-control) (Mas-9th-5 min means the Raman spectra after 9th stretched load and unload for 5 min and so on)
表1 蜘蛛大壶状丝拉曼光谱在反复拉伸过程中特征谱带及其波谱数的变化
Table 1
| Control | 9th- 5 min | 9th- 2 h | 9th- 20 h | 14th- 15 min | 23rd- 5 min | 27th- 5 min | 34th- 5 min | Assignment[30, 31] |
|---|---|---|---|---|---|---|---|---|
| 1693 | 1697 | 1697 | - | 1697 | 1697 | 1694 | 1697* | Amide Ⅰ, C=O in antiparallel β-sheet |
| 1678 | - | - | 1671 | 1669 | 1672 | 1669 | 1669 | Amide Ⅰ, C=O in β-sheet or random coil |
| - | 1655* | 1655 | - | 1651 | - | - | - | Amide Ⅰ, α-helix or random coil poly(E) |
1636 1615 - | 1636 1615 1593 | 1636 1613 1593 | 1636 1613 - | 1637 1611 1595 | 1636 1615 1597* | 1636 1615 1594 | 1633 1610 1594 | Amide Ⅰ, β-sheet aromatic ring s in F and Y |
| 1560 | 1558 | 1567 | 1567 | 1555/1545 | 1567 | 1567 | 1562 | Amide Ⅱ, C-N α-helix or random coil |
| 1523 | 1523 | 1523 | 1523 | 1523 | 1523 | 1523 | 1522 | Amide Ⅱ, β-sheet |
| 1449 | 1449 | 1449 | 1443 | 1452 | 1445 | 1445 | 1445 | Poly(A) CH3 ab, poly(AG) CH2 b |
| - | 1440 | - | - | 1442 | - | - | - | COO- ss in D and E |
1386 1362 | 1386 1362 | 1390 1362 | 1386 1362 | 1385 1362 | 1386 1361 | 1386 1361 | 1381 1360 | Amide Ш, Ha b, N-H ib |
| 1335 | 1335 | 1328 | 1324 | 1325 | 1325 | 1325 | 1318 | Amide Ш, CH3 sb, Ha b |
| 1277 | 1277 | 1277 | 1277 | 1277 | 1277 | 1277 | 1276 | Amide III: N-H ib, C=O ib, C-C s |
| 1243 | 1237 | - | 1227 | 1224 | 1219 | 1220 | Amide Ш, N-H ib, CH2 tw | |
| - | 1186 | - | - | 1186 | - | - | - | Amide Ш, aromatic ring C-H ib |
| - | - | - | 1147 | 1147 | - | - | - | H a b |
| 1117 | 1123 | 1123 | 1123 | 1123 | 1123 | 1123 | - | C-C β s |
| 1096 | 1096 | 1096 | 1096 | 1096 | 1095 | 1095 | 1095 | C-C β s |
| 1045 | 1045 | 1045 | 1045 | 1046 | 1045 | 1045 | 1039 | C-C β s |
| - | 1002 | 1003 | 1003 | 1004 | 1002 | 1002 | 997 | Aromatic ring br in F and Y |
968 924 904 | 951 924 904 | 962 924 904 | 962 924 904 | 947 926 904 | 947 918* 906 | 948 926* 906 | 946 - - | Poly-alanine CH3r, N-C α s, C-C s, C=O s |
3 讨论
3.1 蜘蛛大壶状腺丝的形变、力学行为与蛋白二级结构之间的关系
抗拉伸疲劳的能力是衡量纤维材料性能的一个重要参数[32]。在纤维材料的制备过程中,部分程序或工艺是使纤维处于被反复定向拉伸处理,以诱导其分子定向结晶、提高拉伸方向的强度[33]。而在自然条件下大壶状腺丝也会被反复拉伸,但反复拉伸对蛛丝的结构和力学性能的影响缺少深入系统的研究[18~21]。大壶状腺丝纺出后,随着放置时间的增加其力学性能略有提高[13]。本研究结果表明,蜘蛛大壶状腺丝能反复定伸长拉伸几十次至上百次,具有良好的反复拉伸特性与抗拉伸疲劳的能力。蜘蛛大壶状腺丝反复拉伸过程中其形变、力学行为与蛋白二级结构之间的关系列于表2:(1)每次拉伸前蛛丝原长变化差异不显著(P>0.05),力学行为曲线重叠性好(图5a、b、e、f、k、l;图6a~f,图7e~j、m、n和图9g、h、k、l),表明蛛丝的蛋白二级结构没有发生变化;(2)每次拉伸前蛛丝原长变化差异不显著(P>0.05),力学行为曲线的重叠性较差(图6g~j和图7o、p),表明其蛋白二级结构发生了变化;(3)每次拉伸前蛛丝原长变化差异显著(P<0.05),力学行为曲线重叠性好(图5c、d、g~j,图7a~d和图9e),表明其蛋白二级结构发生了精确的变化;(4)每次拉伸前蛛丝原长变化差异显著(P<0.05),力学行为曲线重叠性差,表明其蛋白二级结构可能发生了某种更大的跃迁性变化(图6k、l和图7k、l)。
表2 蜘蛛大壶状腺丝的形变、力学行为与蛋白二级结构之间的关系
Table 2
| Geometric parameter | Statistical hypothesis test | Tensile behavior curve | |
|---|---|---|---|
| Repeatability (good) | Repeatability (bad) | ||
| Initial length (L0) | Unchanged (P>0.05) | Structures unchanged | Structures change |
| Changed (P<0.05) | Structures precise change | Structures change | |
蜘蛛大壶状腺丝在反复拉伸过程中,在定伸长长度不变或增加很少的情况下,蛛丝的原长变化差异不显著(P>0.05),即抗拒被拉长和抗拒形变。两次拉伸之间的间隔相同时间蛛丝蛋白二级结构不变,力学行为曲线重叠性好(图5a、b、e、f、k、l,图6a~f,图7e~j、m、n和图9g、h、k、l),或蛛丝蛋白二级结构发生了改变,力学行为曲线重叠性较差(图6g~j和图7o、p)。蛛丝抗拒拉长和抗拒形变,还表现在组间长时间间隔后蛛丝拉伸前的原长较之前变短(P<0.05)。纤维材料的形状记忆与其力学行为的关系密切[34]。Emile等[35]研究表明,大壶状腺丝具有横向扭曲的形状记忆能力;而本研究表明,大壶状丝具有纵向的形状记忆能力,即蜘蛛大壶状腺丝不论是横向扭曲还是纵向拉伸都具有形变记忆能力。定伸长长度增加幅度继续增加,蛛丝无法抗拒被拉长即发生了较小的形变,两次拉伸间隔相同的时间,蛛丝蛋白二结构发生了精确的变化,因此前后的力学行为曲线重叠性好;定伸长长度继续较大幅度地增加,蛛丝最终被拉长而发生了较大的形变,两次拉伸之间间隔相同的时间而且蛛丝结构发生了跃迁性变化,如同上了另外一个结构台阶(图6k、l;图7k、l)。在同一层台阶,蛛丝倾向于保持其原长,即表现出纵向的形状记忆。如果原长发生了较小但是差异显著的变化,即发生了较小的形变,则蛛丝蛋白二级结构会发生精确的变化,力学行为曲线仍然重叠得很好(图5c、d、g~j和图7a~d)。但是,这种变化不足以使蛛丝跨上一个新结构台阶。这似乎表明,蛛丝的形变、力学行为与蛋白二级结构之间存在某种映射关系。
前后两次拉伸间隔不同时间梯度的力学行为曲线,表现出一部分不受拉伸历史的影响而重现之前的力学行为,另一部分则不能重现之前的力学行为(图8)。这可以解释为,拉伸后蛛丝蛋白二级结构的恢复可能有一个间隔时间最小阈值和最大阈值,即间隔时间达到最小阈值蛛丝蛋白二级结构才能恢复,间隔时间一旦达到最大阈值无论间隔再增加多少时间蛛丝蛋白二级结构都不发生改变,而上一次拉伸蛛丝蛋白二级结构一旦发生了跃迁性变化,则无论间隔多少时间下次拉伸都不能重现之前的力学行为。这似乎表明,蜘蛛丝具有一个有记忆的“结构时钟”。蜘蛛丝表现出一种良好的力学行为记忆能力,即前后两组拉伸之间长时间(≥23 min)间隔后只需经过一次拉伸就能不受拉伸历史的影响重现之前的力学行为(图7a~j、m、n和图9g、h、k、l)。有时在伸长长度增加的情况下也只需要一次拉伸随后蛛丝的力学行为曲线也重叠的很好(图5c、d、g、h、k、l和图6a、b),表现出类似于橡胶的力学行为特征[36]。这不符合材料学中塑性形变不可逆的普遍一般原则。此外,本文还发现大壶状腺丝在定伸长逐渐增加反复拉伸至一定次数后,其力学性能趋于稳定,达到一种最佳状态。随着拉伸次数与定伸长长度的增加其力学行为的屈服点负荷逐渐下降,但是其屈服强度基本上保持不变(图2、图6k、l和图9a~d);另外,有的拉伸组的位移-负载曲线其加强区有小幅度发散,但是其应力-应变曲线又聚集在一起表现出比位移-负载曲线更好的重叠性(图5c、d、g、h和图7e、f、k、l)。这似乎表明,蛛丝可以通过几何尺寸调节其力学性能的再现性,具有一种良好的力学性能后加工调节能力。
3.2 反复拉伸对蜘蛛大壶状腺丝蛋白二级结构的影响
酰胺Ⅰ(约1650 cm-1)和酰胺Ⅱ(约1540 cm-1)的峰是无序结构的贡献,表明可能含有自由卷曲和/或α-螺旋结构。这些结构形成无定型非结晶区,使丝纤维具有延展性;酰胺Ⅰ(约1620 cm-1)和酰胺Ⅱ(约1529 cm-1)的峰则与β-折叠有关,酰胺Ⅰ(约1669 cm-1)附近的吸收峰可能是与β-折叠结构有关的β-转角或β-弯曲的吸收峰。这些结构形成结晶区,使丝纤维具有高模量和高强力[37, 38]。Yasumasa Fukushima[37]设计了以X-Gly-Gly(X分别为Ala,Ty,Gln和Leu)为基序的4个肽链分子,作为蜘蛛纺自大壶状腺的拖牵丝富含Gly序列的模型。4种聚合分子的红外光谱表明,随着分子中β-折叠含量的提高Poly(Leu-Gly-Gly)<Poly(Ala-Gly-Gly)<Poly(Gln-Gly-Gly)=Poly(Tyr-Gly-Gly),在波谱数约为1695、1520和1626 cm-1处的吸收峰随之增强;随着分子中无序结构的增加,酰胺Ⅰ(约1650 cm-1)和酰胺Ⅱ(约1540 cm-1) 的吸收峰增加。在800~1110 cm-1发生的形变属于β-折叠,960 cm-1左右为PolyAla特征峰[30]。以此为依据,本文研究的大壶状腺丝在第1次拉伸前的拉曼光谱中出现了表征自由卷曲和/或α-螺旋结构的酰胺Ⅰ(约1640 cm-1)和酰胺Ⅱ(约1560 cm-1)特征谱带,也出现了表征β-折叠或者β-转角或者β-弯曲的酰胺Ⅰ(约1615/1678 cm-1)、酰胺Ⅱ(约1523 cm-1)和PolyAla(约968 cm-1)的特征谱带(图10a Mas-0-control)。这些特征谱带与文献报道的结果基本一致,表明蜘蛛大壶状腺丝是一种半结晶的天然动物蛋白纤维。
与第一次拉伸前的对照相比,蛛丝在被第9次定伸长反复拉伸过屈服点屈服区至加强区松弛5 min后,其拉曼光谱酰胺Ⅰ(约1678 cm-1)特征谱带消失,酰胺Ⅱ(约1560 cm-1)特征谱带偏移至1558 cm-1且峰强度降低,同时出现新的表征β-折叠的酰胺Ⅰ(约1593 cm-1),而原来的1117 cm-1转移到1123 cm-1,说明在拉伸后发生了C-C键的形变[30],并经20 h后无法恢复,表明β-折叠又重新形成;被多次拉伸后新出现了1147和1186 cm-1两个峰,1243 cm-1偏移到1237 cm-1,这些都属于β-折叠的C-C对称结构的形变[32],表明原有的β-折叠结构已经发生了结构形变,同时形成了新的PGII型β-折叠结构(图11);第9次拉伸结束松弛之后2 h的拉曼光谱其酰胺Ⅱ(约1567 cm-1)特征谱带又重新开始出现,同时新出现了酰胺Ⅰ(约1655 cm-1),这表明有一部分又重新回复成了自由卷曲结构;第9次拉伸结束松弛之后20 h的拉曼光谱其酰胺Ⅱ(约1567 cm-1)特征谱带继续增强基本回到第一次拉伸之前的强度,1560 cm-1拉伸后消失,随着时间的推移又出现,并变成1567 cm-1附近,可能属于少量色氨酸残基,拉伸后可能被遮盖,随后又暴露出来[31],而之前新出现的酰胺Ⅰ(约1655 cm-1)消失,同时酰胺Ⅰ(约1671 cm-1)出现,即经过过夜长时间间隔后使整个光谱基本恢复到第1次拉伸之前的状态,这与第1次第10次的拉伸力学行为曲线基本相似是一致的。第14次拉伸结束松弛之后15 min的拉曼光谱又出现了表征β-折叠以及可能表征β-转角或者β-弯曲的1669 cm-1特征带、表征β-折叠的酰胺Ⅰ(约1594 cm-1)与表征自由卷曲或者α-螺旋结构的酰胺Ⅰ(约1651 cm-1),表明该结构只需较短时间就能恢复,但酰胺Ш由1335 cm-1偏移至1325 cm-1左右经过15 min甚至过夜都无法恢复,表明已有形变发生。1325 cm-1峰高增强,说明经过多次拉伸后,暴露出α-螺旋[31]。拉伸后出现约1440 cm-1的特征带,开始会随着间隔时间增加而消失,随后第14次拉伸再次出现,可能属于Asp和Glu残基C=O的伸缩振动,说明拉伸后暴露出包含在内部的Asp和Glu[30]。表明多次拉伸后主要是无规则卷曲发生形变,有部分参与形成类似新的β-折叠,有的β-折叠已经不能重新形成,蜘蛛丝的蛋白二级结构如原来的β-折叠已经开始受到破坏。
图11
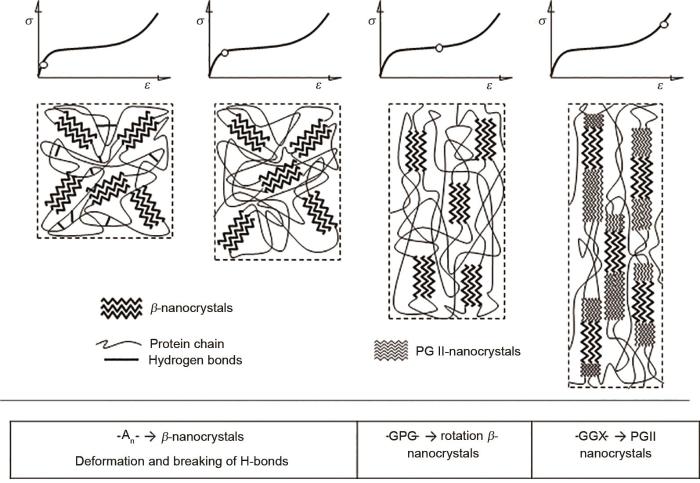
图11
蜘蛛大壶状腺丝拉伸过程中蛋白二级结构的变化模型图
Fig.11
Change model of protein secondary structure during repeated stretching of spider Mas
蛛丝被拉断成为单丝后,第23次拉伸开始,1046 cm-1偏移到1039 cm-1,对照样品的1117 cm-1转移到1123 cm-1,说明C-C伸缩振动在拉伸后发生了形变,并在最后一次拉断后无法恢复,单丝的β-折叠已经再难恢复成原样。1325 cm-1形变剧烈,说明经过多次拉伸后,暴露出α-螺旋[31];PolyAla特征带随着拉伸次数增加其波谱数由第1次拉伸前的968 cm-1偏移至946 cm-1附近,而且PolyAla特征带924 cm-1和904 cm-1早期拉伸后经过时间间隔都有恢复,但被拉断成单股后,这些特征峰开始减弱并在最后一次拉断后消失了,表明原本的二级结构无规则卷曲和β-折叠遭到破坏后无法形成新的β-折叠。蛛丝在被反复拉伸后有些特征谱带波谱数发生了细小的偏移,这与文献报道[39]的大壶状腺丝被拉伸至应变18%时部分特征谱带波谱数发生小的偏移的结果是相一致的。
3.3 蜘蛛丝力学性能策略调节方式
已有的研究结果表明,可调节蛛丝的力学性能的方式有两种,以实现不同的力学性能策略[22]。第一种为改变蛛丝的分子组成和结构,如纺自大壶状腺的卵袋框丝与纺自柱状腺的卵袋包卵丝的氨基酸组成与蛋白二级结构有很大的不同[23, 24]。即使同一丝腺纺出的丝纤维的氨基酸组成与蛋白结构也有不同[40, 41],如大壶状腺至少含有两种上皮细胞,分别分泌MaSp1与MaSp2(大壶状腺丝心蛋白1与2)这两种丝心蛋白家族成员,MaSp1不含有脯氨酸,MaSp2富含脯氨酸。当MaSp2流经大壶状腺膨大区时,MaSp1分泌出与MaSp2以不同比例混合流经导管至纺管最后被纺出[41]。第二种方式为改变丝纤维的几何尺寸如原长、直径等[27]。本研究结果表明,蛛丝不但可通过蛋白二级结构和几何尺寸的变化,还可通过拉伸后间隔的时间效应调节其力学行为与性能,表现出力学行为记忆能力,即前后两组相同短时间间隔(≤5 min)的拉伸之间经过长时间间隔(≥23 min)后蛛丝只需一次拉伸就可重现之前的力学行为。这是蜘蛛大壶状腺丝的一种新的力学性能策略。
本文设计的实验较好的模拟了大壶状腺丝在自然界中被反复拉伸的情形,也验证了我们的推测:蜘蛛大壶状腺丝反复拉伸特性是蜘蛛对其丝纤维力学性能的一种后加工调节方式,表现为大壶状腺丝纺出后通过反复拉伸改变其结构以调节定制(Tailor)其力学性能。
4 结论
(1) 蜘蛛大壶状腺丝能反复定伸长拉伸几十次至上百次,且越拉越硬,但其屈服强度基本保持不变,表明其具有良好的反复拉伸特性与抗拉伸疲劳的能力。
(2) 蜘蛛大壶状腺丝具有良好的力学行为记忆能力,即前后两组拉伸之间长时间(≥23 min)间隔后只需经过一次拉伸就能不受拉伸历史的影响重现之前的力学行为。这不符合材料学中塑性形变不可逆的普遍一般原则,表现出类似于橡胶的力学行为特征。这种力学行为记忆能力可通过两种方式实现,一是保持不发生形变(即纵向的形状记忆)与蛋白二级结构的变化,二是发生了形变但蛋白二级结构发生了精确的变化。
(3) 蜘蛛大壶状腺丝表现出一种新的力学性能策略:力学行为的记忆即保持力学行为不变,在反复拉伸后以及两次拉伸之间间隔不同的时间会使蛛丝蛋白二级结构如自由卷曲中的氢键断裂,也会重新形成β-转角或β-弯曲结构以及PGⅡ β-折叠,以此来调节蛛丝蛋白二级结构以实现其力学行为的记忆。
(4) 蜘蛛大壶状腺丝力学行为虽然具有弹性区、屈服点、屈服区和加强区,但是整体表现出类似于橡胶材料的超弹性性能。
致谢
感谢复旦大学高分子科学系孙畅与杨占峰实验师对本文蛛丝在反复拉伸过程中蛋白二级结构变化的研究提供的帮助,感谢西南民族大学青藏高原研究院2020级硕士研究生唐思思以及湖南师范大学生命科学学院2020级硕士研究生陈颖对本论文所做的一小部分研究工作。
参考文献
Duplication and concerted evolution of MiSp-encoding genes underlie the material properties of minor ampullate silks of cobweb weaving spiders
[J].
Background: Orb-web weaving spiders and their relatives use multiple types of task-specific silks. The majority of spider silk studies have focused on the ultra-tough dragline silk synthesized in major ampullate glands, but other silk types have impressive material properties. For instance, minor ampullate silks of orb-web weaving spiders are as tough as draglines, due to their higher extensibility despite lower strength. Differences in material properties between silk types result from differences in their component proteins, particularly members of the spidroin (spider fibroin) gene family. However, the extent to which variation in material properties within a single silk type can be explained by variation in spidroin sequences is unknown. Here, we compare the minor ampullate spidroins (MiSp) of orb-weavers and cobweb weavers. Orb-web weavers use minor ampullate silk to form the auxiliary spiral of the orb-web while cobweb weavers use it to wrap prey, suggesting that selection pressures on minor ampullate spidroins (MiSp) may differ between the two groups.Results: We report complete or nearly complete MiSp sequences from five cobweb weaving spider species and measure material properties of minor ampullate silks in a subset of these species. We also compare MiSp sequences and silk properties of our cobweb weavers to published data for orb-web weavers. We demonstrate that all our cobweb weavers possess multiple MiSp loci and that one locus is more highly expressed in at least two species. We also find that the proportion of beta-spiral-forming amino acid motifs in MiSp positively correlates with minor ampullate silk extensibility across orb-web and cobweb weavers.Conclusions: MiSp sequences vary dramatically within and among spider species, and have likely been subject to multiple rounds of gene duplication and concerted evolution, which have contributed to the diverse material properties of minor ampullate silks. Our sequences also provide templates for recombinant silk proteins with tailored properties.
Almost a spider: a 305-million-year-old fossil arachnid and spider origins
[J].
Material properties of evolutionary diverse spider silks described by variation in a single structural Parameter
[J].Spider major ampullate gland silks (MAS) vary greatly in material properties among species but, this variation is shown here to be confined to evolutionary shifts along a single universal performance trajectory. This reveals an underlying design principle that is maintained across large changes in both spider ecology and silk chemistry. Persistence of this design principle becomes apparent after the material properties are defined relative to the true alignment parameter, which describes the orientation and stretching of the protein chains in the silk fiber. Our results show that the mechanical behavior of all Entelegynae major ampullate silk fibers, under any conditions, are described by this single parameter that connects the sequential action of three deformation micromechanisms during stretching: stressing of protein-protein hydrogen bonds, rotation of the beta-nanocrystals and growth of the ordered fraction. Conservation of these traits for over 230 million years is an indication of the optimal design of the material and gives valuable clues for the production of biomimetic counterparts based on major ampullate spider silk.
Sequential origin in the high performance properties of orb spider dragline silk
[J].Major ampullate (MA) dragline silk supports spider orb webs, combining strength and extensibility in the toughest biomaterial. MA silk evolved ~376 MYA and identifying how evolutionary changes in proteins influenced silk mechanics is crucial for biomimetics, but is hindered by high spinning plasticity. We use supercontraction to remove that variation and characterize MA silk across the spider phylogeny. We show that mechanical performance is conserved within, but divergent among, major lineages, evolving in correlation with discrete changes in proteins. Early MA silk tensile strength improved rapidly with the origin of GGX amino acid motifs and increased repetitiveness. Tensile strength then maximized in basal entelegyne spiders, ~230 MYA. Toughness subsequently improved through increased extensibility within orb spiders, coupled with the origin of a novel protein (MaSp2). Key changes in MA silk proteins therefore correlate with the sequential evolution high performance orb spider silk and could aid design of biomimetic fibers.
Spider dragline silk: correlated and mosaic evolution in high-performance biological materials
[J].The evolution of biological materials is a critical, yet poorly understood, component in the generation of biodiversity. For example, the diversification of spiders is correlated with evolutionary changes in the way they use silk, and the material properties of these fibers, such as strength, toughness, extensibility, and stiffness, have profound effects on ecological function. Here, we examine the evolution of the material properties of dragline silk across a phylogenetically diverse sample of species in the Araneomorphae (true spiders). The silks we studied are generally stronger than other biological materials and tougher than most biological or man-made fibers, but their material properties are highly variable; for example, strength and toughness vary more than fourfold among the 21 species we investigated. Furthermore, associations between different properties are complex. Some traits, such as strength and extensibility, seem to evolve independently and show no evidence of correlation or trade-off across species, even though trade-offs between these properties are observed within species. Material properties retain different levels of phylogenetic signal, suggesting that traits such as extensibility and toughness may be subject to different types or intensities of selection in several spider lineages. The picture that emerges is complex, with a mosaic pattern of trait evolution producing a diverse set of materials across spider species. These results show that the properties of biological materials are the target of selection, and that these changes can produce evolutionarily and ecologically important diversity.
The strucure and properties of spider silk
[J].
Strength and structure of spider' silks
[J].
Composition, structure and biological function of spider silk
[J].
蜘蛛丝的组成结构与生物学功能
[J].
Multiscale mechanisms of nutritionally induced property variation in spider silks
[J].
Physicochemical Property Variation in Spider Silk: Ecology, Evolution, and Synthetic Production
[J].The unique combination of great stiffness, strength, and extensibility makes spider major ampullate (MA) silk desirable for various biomimetic and synthetic applications. Intensive research on the genetics, biochemistry, and biomechanics of this material has facilitated a thorough understanding of its properties at various levels. Nevertheless, methods such as cloning, recombination, and electrospinning have not successfully produced materials with properties as impressive as those of spider silk. It is nevertheless becoming clear that silk properties are a consequence of whole-organism interactions with the environment in addition to genetic expression, gland biochemistry, and spinning processes. Here we assimilate the research done and assess the techniques used to determine distinct forms of spider silk chemical and physical property variability. We suggest that more research should focus on testing hypotheses that explain spider silk property variations in ecological and evolutionary contexts.
variability in the mechanical properties of spider silks on three leveles: interspecific, intraspecific and intraindividual
[J].The mechanical characteristics of dragline silks collected from a range of spiders drawn from the Argiopidae, Tetragnathidae, Theridiidae and Pisauridae displayed significant inter- and intraspecific differences. Dragline silks of the same species could show considerable variability probably dependent upon spider condition: starvation, for example, lead to decreased breaking elongation in Nephila edulis. Environmental conditions such as reeling speed affected silk properties such that (i) breaking elongation decreased, (ii) breaking stress increased and (iii) Young's modulus increased with increasing reeling speed. However, N. edulis and Araneus diadematus responded differently to the reeling speed treatments suggesting differences in basic silk properties.
The effects of seasonal changes on the molecular weight of Nephila clavata spider silk
[J].
Finding inspiration in argiope trifasciata spider silk fibers
[J].
Insights into silk formation process: correlation of mechanical properties and structural evolution during artificial spinning of silk fibers
[J].
Reconfigurable bioinspired framework nucleic acid nanoplatform dynamically manipulated in living cells for subcellular imaging
[J].
Biomimetic supertough and strong biodegradable polymeric materials with improved thermal properties and excellent uv-blocking performance
[J].
The science of silks
[J].
Analysis of spider silk in native and supercontracted states using Raman spectroscopy
[J].
Relationship of Tensile behaviors and biological function of silk fibers from egg case of argiope bruennichi
[J].
横纹金蛛卵袋丝的力学行为与生物学功能之间的关系
[J].
Recovery in spider silk fibers
[J].
Recovery in Viscid Line Fibers
[J].
Morphologies and tensile behaviors of three types of spider silks with different functions
[J].
三种不同功能蛛丝的超微结构与拉伸力学行为
[J].
Physico-chemical structural characterizations and mechanical properties of egg case silk from the two spiders: Argiope Amoena and Nephila Clavata
[J].
悦目金蛛和棒络新妇卵袋丝物理化学结构表征及其力学性能
[J].
Amino acid profiles and tensile behaviors of egg case silk of spider Nephila clavata
[J].
棒络新妇卵袋丝氨基酸组成及其力学行为
[J].
Spider silk gut: development and characterization of a novel strong spider silk fiber
[J].Spider silk fibers were produced through an alternative processing route that differs widely from natural spinning. The process follows a procedure traditionally used to obtain fibers directly from the glands of silkworms and requires exposure to an acid environment and subsequent stretching. The microstructure and mechanical behavior of the so-called spider silk gut fibers can be tailored to concur with those observed in naturally spun spider silk, except for effects related with the much larger cross-sectional area of the former. In particular spider silk gut has a proper ground state to which the material can revert independently from its previous loading history by supercontraction. A larger cross-sectional area implies that spider silk gut outperforms the natural material in terms of the loads that the fiber can sustain. This property suggests that it could substitute conventional spider silk fibers in some intended uses, such as sutures and scaffolds in tissue engineering.
Preparation and characterization of Nephila clavipes tubuliform silk gut
[J].
Tensile behavior and morphology of differently degummed Silkworm cocoon silk fibres
[J].
Volume constancy during stretching of spider silk
[J].The characterization of silk properties requires a reliable measurement of stress-strain curves from tensile tests, which calls for a detailed analysis of what is considered the cross section of the sample and how it varies during the experiments. Here, spider silk fibers from the major ampullate gland (MAS) of Argiope trifasciata spiders are tensile tested, and the cross-sectional area is measured under different strained configurations. It has been found that the fiber volume remains practically constant during stretching, and deformation proceeds homogeneously in all the fibers. The conservation of volume is validated independently of the type of fiber and the strain level. This result, applied to compute true stress-strain curves for different MAS fibers, shows that the description of their properties depends noticeably on which set of tensile parameters is chosen (true or engineering), and that engineering values could lead to misinterpretation of experiments that combine results from different strain ranges.
Stretching of supercontracted fibers: a link between spinning and the variability of spider silk
[J].
Study of protein conformation and orientation in silkworm and spider silk fibers using raman microspectroscopy
[J].
Raman spectroscopy of proteins
[J].
Analysis of fatigue test parameters of nylon warp knitted fabric
[J].
氯纶经编针织物疲劳性试验参数的分析
[J].
Preparation and characterization of polymethylmetharylate/aligned SWCNT composites with by repeated stretching
[J].
反复拉伸法制备单壁碳纳米管定向排列的SWCNT/PMMA复合材料
[J].
Mechanic behaviors of the thermoviscoelastic matrix composites with shape memory fiber
[J].
形状记忆纤维热粘弹性基体复合材料的力学行为
[J].
Shape memory in spider dragline
[J].
Mechanical behavior and stretching crystallization of rubber
[J].
橡胶的力学行为和拉伸结晶化
[J].
Secondary structural analysis in the solid state for analogous sequential polypeptides of glycine-rich sequence of spider dragline silk
[J].
Conformation studies of silk proteins with infrared and raman spectroscopy
[J].
红外和拉曼光谱用于对丝蛋白构象的研究
[J].
Molecular deformation in spider dragline silk subjected to stress
[J].
Protein and amino acid composition of silks from the cob weaver, Latrodectus Hesperus (black widow)
[J].The silks from the cob weaving spider, Latrodectus hesperus (black widow), have been examined with the goal of expanding our understanding of the relationship between the protein structure and mechanical performance of these unique biomaterials. The scaffolding, dragline and inner egg case silks each appear to be distinct fibers based on mole percent amino acid composition and polypeptide composition. Further, we find that the amino acid composition of dragline and egg case silk are similar to the analogous silks produced by orb weaving spiders, while scaffolding silk may represent a novel silk. The black widow silks are comprised of multiple high molecular weight polypeptides, however, the egg case and scaffolding silks also contain some smaller polypeptides.
Spider silk protein refolding is controlled by changing pH
[J].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}